Fr, 29.07.2016 - 07:49 — Christian Körner

![]()
Unsere belebte Welt basiert auf dem Element Kohlenstoff. Anders als alle anderen Elemente kann Kohlenstoff auf Grund seiner einzigartigen Chemie eine ungeheure Mannigfaltigkeit an Biomolekülen bilden und bleibt darin auch die Hauptkomponente. Dementsprechend besteht die gesamte Biomasse (bezogen auf ihr Trockengewicht) zu rund 50 % aus Kohlenstoff.
Der Kohlenstoff durchläuft einen globalen Kreislauf; für die Biosphäre sind darin zwei grundlegende Funktionen bestimmend:
- die Input-Funktion - die Aufnahme (Assimilation) von Kohlenstoff in seiner oxydierten Form CO2 aus der Luft durch die Ökosysteme der Pflanzenwelt via Photosynthese - und
- die Output Funktion, d.i. der Prozess der Atmung, über den alle Organismen CO2 an die Atmosphäre wieder abgeben (Abbildung 1):
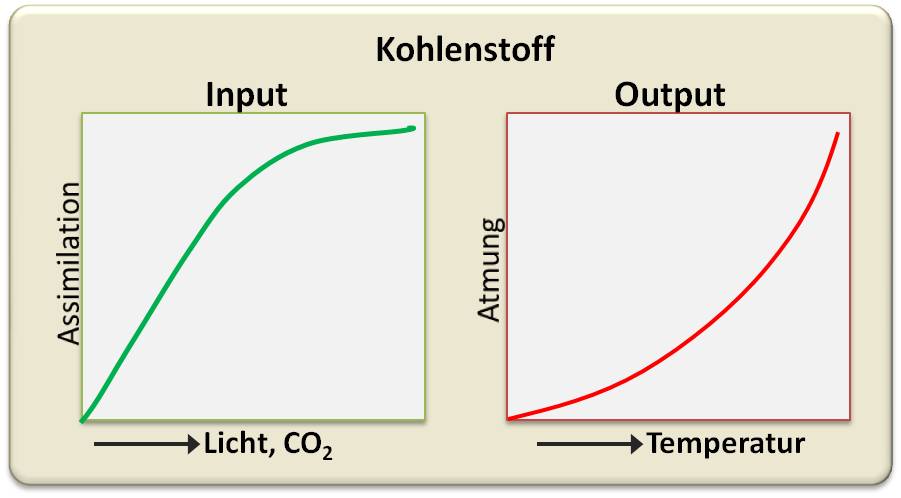 Abbildung 1. Input von CO2 via Photosynthese und Output via Atmung sind grundlegende Funktionen im globalen Kohlenstoffkreislauf.
Abbildung 1. Input von CO2 via Photosynthese und Output via Atmung sind grundlegende Funktionen im globalen Kohlenstoffkreislauf.
Die Input-Funktion (grüne Kurve in Abbildung 1) zeigt, dass die Kohlenstoffaufnahme von vorhandenem Licht und CO2 abhängt und steigt, wenn diese zunehmen, schließlich aber in eine Sättigung übergeht . Die rote Kurve gibt eine grundsätzliche Charakteristik physiologischer Vorgänge wieder, dass nämlich (bio)chemische Prozesse - die Atmung mit eingeschlossen - mit steigender Temperatur schneller werden und damit auch wieder mehr CO2 freigesetzt wird.
An die mit zunehmendem CO2 steigende Inputfunktion knüpft sich eine weitverbreitete Hoffnung, dass mehr CO2 in der Atmosphäre auch ein Mehr an Pflanzenwuchs und damit einen Vorteil für das Leben auf unserem Planeten bewirken sollte.
Wodurch wird das Pflanzenwachstum gesteuert?
Das Pflanzenwachstum nur als Funktion von Input und Output zu behandeln, ist zweifellos zu kurz gegriffen. Ob der Kohlenstoffpool in der Biosphäre steigt oder sinkt wird tatsächlich durch die Wechselwirkungen zwischen Input- und Output bestimmt und diese hängen von Umweltbedingungen und verfügbaren Ressourcen ab. Ausreichend Licht vorausgesetzt sind dies
ausreichend Wasser, adäquate Temperaturen und das Vorhandensein von Nährstoffen.
Für die Existenz von Leben sind außer Kohlenstoff ja noch 24 weitere chemische Elemente essentiell. Während Kohlenstoff und Stickstoff in der Atmosphäre theoretisch unbegrenzt für den Bedarf der Pflanzenwelt zur Verfügung stehen, ist das Vorkommen anderer Ressourcen, wie etwa das von Phosphor, Kalium, Magnesium, Mangan, Molybdän, Selen, u.a. limitiert und von der Gegend und dem jeweiligen Boden abhängig.
Pflanzenwachstum als Baustelle betrachtet
Wenn man ein Haus bauen will, braucht man natürlich Bausteine, die in einer Fabrik (ich benutze dafür den Term "Quelle") hergestellt werden, und Maurer, die das Haus bauen. Die Schnelligkeit mit welcher an der Baustelle (für die ich den Term "Senke" verwende) der Hausbau erfolgt, hängt von vielen Umständen ab: vom Arbeitseinsatz der Maurer, vom Vorhandensein benötigter Materialien, von den Wetterbedingungen, etc. - zweifellos aber kaum von der Zustellrate der Bausteine zur Baustelle. Diese erfolgt dem Baufortschritt entsprechend, nach Bedarf. Abbildung 2.
 Abbildung 2. Das Wachstum einer Pflanze gleicht dem Bau eines Hauses. Ausgangsstoffe werden in der Photosynthese/Fabrik - d.i. an der Quelle – in Baumaterial (Zucker, Ziegel) umgewandelt, an die Baustelle geschafft und dort (von Biokatalysatoren, Maurern) zum Aufbau des Baumes/Hauses verwendet. Wie schnell an dieser Senke dann der Aufbau erfolgt, wird zumeist durch limitierende "Umweltbedingungen" und nur selten durch den Antransport der Baustoffe bestimmt (aus C. Körner (2012) Biologie in unserer Zeit 42:238-243)
Abbildung 2. Das Wachstum einer Pflanze gleicht dem Bau eines Hauses. Ausgangsstoffe werden in der Photosynthese/Fabrik - d.i. an der Quelle – in Baumaterial (Zucker, Ziegel) umgewandelt, an die Baustelle geschafft und dort (von Biokatalysatoren, Maurern) zum Aufbau des Baumes/Hauses verwendet. Wie schnell an dieser Senke dann der Aufbau erfolgt, wird zumeist durch limitierende "Umweltbedingungen" und nur selten durch den Antransport der Baustoffe bestimmt (aus C. Körner (2012) Biologie in unserer Zeit 42:238-243)
Dass Bausteine nach Bedarf zugestellt werden, gilt ebenso für das Pflanzenwachstum, für die Produktivität im Ackerbau und insgesamt für die Produktivität der Biosphäre. Die Fabrik (Quelle) ist hier die Photosynthese, die den Rohstoff des Lebens - Zucker - herstellt und dieser wird dann in das Wachstum der Pflanze investiert. Wie schnell das Wachstum aber erfolgt, ist von der Aktivität der der teilungsfähigen Bildungsgewebe (Meristeme) und damit von der Verfügbarkeit benötigter Ressourcen abhängig. Der Zucker wird nach Bedarf über das Leitungssystem (Phloem) der Pflanze bezogen.
Ein Paradigmenwechsel
Wir alle sind mit der Vorstellung aufgewachsen, dass die Assimilation von CO2 - die Photosynthese - der entscheidende Treiber für das Pflanzenwachstum ist. (Das ist gerade so als ob wir annehmen würden, dass der Baufortschritt eines Hauses von der Produktionsrate der Baustoffe abhängt.) Die Quelle - die Aufnahme von CO2 aus der Atmosphäre - würde die Aktivitäten der Senken - der Stellen, an denen es zur Bildung neuer Zellen, zum Wachstum der Pflanze, zur Bildung neuer Wurzeln, kommt - bestimmen und die Aufnahme anderer benötigter Ressourcen regulieren. Die Versorgung mit CO2 würde also das Wachstum kontrollieren.
Genau das Gegenteil ist der Fall:
Die auf der Verfügbarkeit anderer Ressourcen basierende Aktivität des Meristems zur Gewebebildung steuert je nach Bedarf an Zucker die Photosynthese, reguliert also die Aktivität dieser Quelle. Abbildung 3.
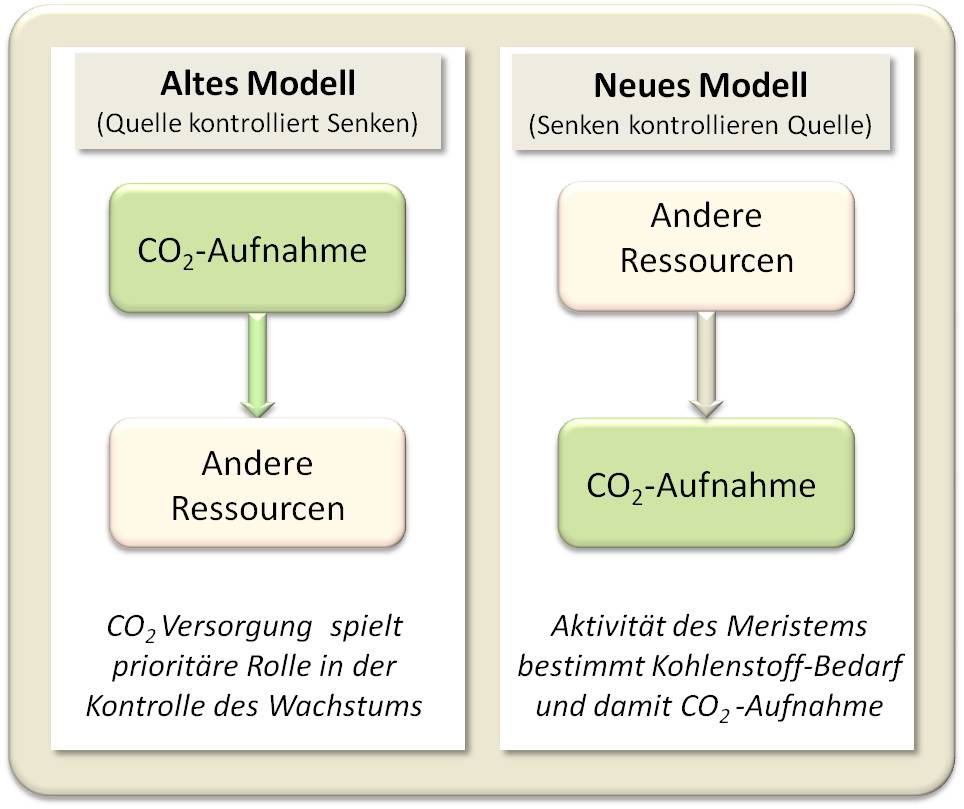 Abbildung 3. Die Aktivität der Gewebebildung (Senke) reguliert den Bedarf an Zucker und damit die CO2-Aufnahme (C. Körner (2015) Curr.Opinion in Plant Biology 25:107-114).
Abbildung 3. Die Aktivität der Gewebebildung (Senke) reguliert den Bedarf an Zucker und damit die CO2-Aufnahme (C. Körner (2015) Curr.Opinion in Plant Biology 25:107-114).
Limitierende Bedingungen
Im allgemeinen wird das Pflanzenwachstum durch den Mangel an anderen Ressourcen als an CO2 begrenzt, das in der Atmosphäre ja reichlich vorhanden ist. Dies möchte ich in der Folge an den limitierenden Bedingungen Trockenheit und Kälte aufzeigen.
Betrachten wir vorerst den Wassermangel,
so kommt es bei zunehmender Trockenheit zu einer raschen Reduzierung des Wachstums und schließlich zu dessen völligem Erliegen. Die Photosynthese wird dagegen erst bei wesentlich höherem Trockenheitsstress verringert. Diese Charakteristik zeigt sich bei allen bis jetzt untersuchten Pflanzensystemen: Wassermangel äußert sich zuerst durch reduzierte Leistungsfähigkeit der Senken, d.i . der Gewebeneubildung und erst viel später in der Beschränkung der CO2-Aufnahme. Abbildung 4.
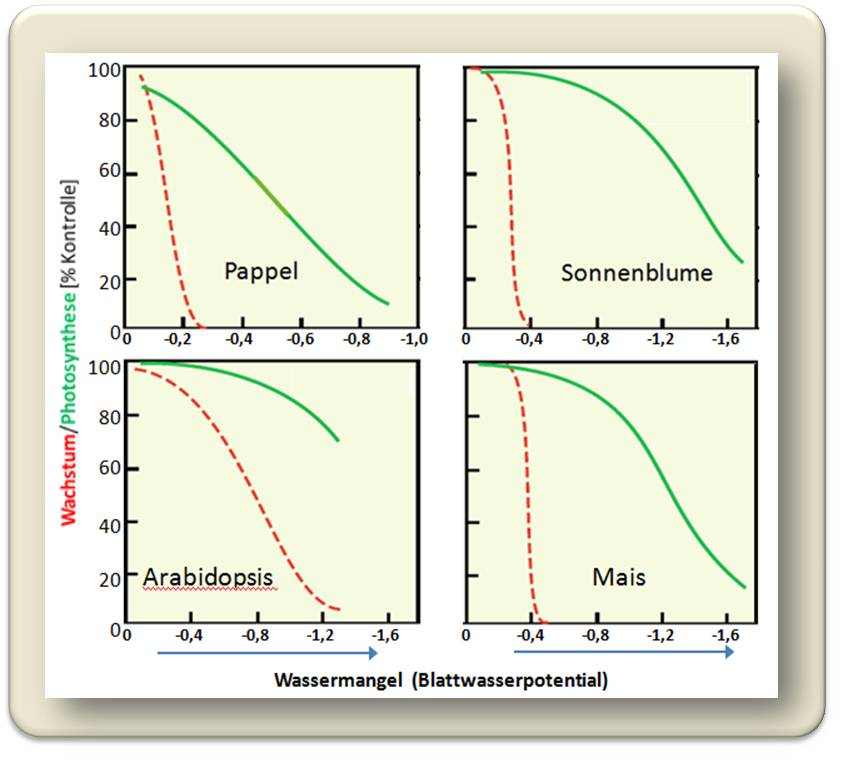 Abbildung 4. Wassermangel bremst primär das Wachstum (rote Kurven) und erst bei viel höherem Wassermangel die Photosynthese(grüne Kurven). Wassermangel ist hier auf das Gewebe (Blattwasserpotential) bezogen und nimmt nach negativen Werten hin zu. (nach Muller et al. 2011, J Exp Bot 62:1715-1729).
Abbildung 4. Wassermangel bremst primär das Wachstum (rote Kurven) und erst bei viel höherem Wassermangel die Photosynthese(grüne Kurven). Wassermangel ist hier auf das Gewebe (Blattwasserpotential) bezogen und nimmt nach negativen Werten hin zu. (nach Muller et al. 2011, J Exp Bot 62:1715-1729).
Für die Folgen von Wassermangel entsteht damit ein neues, völlig anderes Bild:
Nach der alten Lehrmeinung dachte man, dass Blätter bei Trockenheit ihre kleinen Poren ("Stomata") schließen, um dem Wasserverlust vorzubeugen. Dies würde aber um den Preis einer limitierten CO2-Aufnahme erfolgen -somit die Bereitstellung von Zucker durch die Photosynthese- einschränken und als Folge zu einem "Hungern" der Pflanzen führen.
Dieses Bild erweist sich nun als völlig falsch. Tatsächlich geht die Wachstumsreduktion nicht von der Quelle - der Photosynthese - aus, sondern von den Senken: die Produktion neuer Zellen (d.i. vor allem die Bildung der Zellwand aus vornehmlich Zellulose, der auf unserer Erde am häufigsten vorkommenden biochemischen Verbindung) wird stillgelegt, die empfindliche Balance zwischen dem Innendruck (Turgor) der Pflanzenzelle und der Expansion von Zellwand und Zelle gestört. Die Photosynthese läuft indes noch weiter. Es werden Kohlehydrate erzeugt, die allerdings nicht für strukturelles Wachstum eingesetzt werden können, sondern als Zucker und Stärke gespeichert werden. Der Trockenheit ausgesetzte Pflanzen enthalten also tatsächlich mehr und nicht weniger Kohlehydrate, wie die alte Vorstellung von der Hunger leidenden Pflanze glauben machte.
Auch Kälte wirkt sich primär auf die Senken aus
und führt zuerst zur Reduktion des Pflanzenwachstums. Abbildung 5. Zur Verdopplung der Zellzahl - als Maß für das Wachstum genommen - braucht es bei warmer Temperatur rund 10 Stunden, bei Abkühlung auf 10 °C sind es bereits 60 Stunden , bei 5 °C rund 120 Stunden und bei 2 °C geht die Verdopplungszeit gegen unendlich - es findet kein Wachstum mehr statt. Dagegen arbeitet die Photosynthese bereits bei 0 °C mit 30 % und bei 5 °C mit 70 % ihrer maximalen Leistung. Bei weiter steigender Temperatur erreicht die Photosynthese ein breites Maximum zwischen 10 °C und 28 °C und sinkt dann bei noch wärmeren Temperaturen wieder ab.
Dass das Wachstum bei 5 °C aufhört, dürfte für alle Pflanzen gelten, für winterhartes Getreide ebenso wie für andere an Kälte adaptierten Pflanzen und Bäume der Waldgrenze. Welche Mechanismen für diesen universellen Stopp verantwortlich gemacht werden können, ist zur Zeit noch nicht völlig verstanden (es kommen die Proteinsynthese, Energieproduktion in den Mitochondrien, Synthese von Lignin und/oder Zellulose in Betracht). Da die Photosynthese aber noch weiter arbeitet, sollte der Gehalt an niedermolekularen Kohlehydraten in den Pflanzen erwartungsgemäß steigen. Dies wird tatsächlich in der Pflanzenwelt kalter Klimazonen beobachtet.
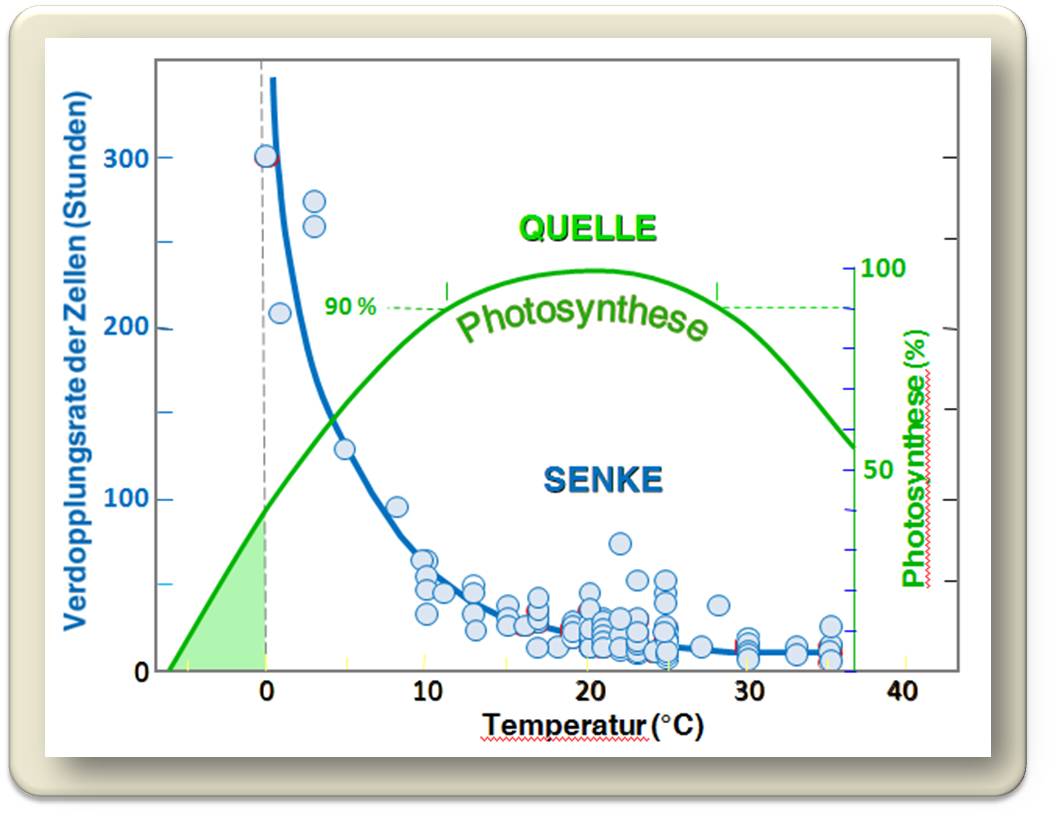 Abbildung 5. Das Pflanzenwachstum wird durch Kälte in viel stärkerem Ausmaß beschränkt als die Photosynthese. Während bei 0 °C kein Wachstum mehr erfolgt, läuft die Photosynthese (grüne Kurve) dagegen noch mit 30 % ihrer maximalen Leistung. Wachstum (blaue Kreise) wurde an Hand der Verdopplungsrate von Kohorten von Zellen gemessen(nach Körner C (2003) Alpine Plant Life. Springer, Berlin).
Abbildung 5. Das Pflanzenwachstum wird durch Kälte in viel stärkerem Ausmaß beschränkt als die Photosynthese. Während bei 0 °C kein Wachstum mehr erfolgt, läuft die Photosynthese (grüne Kurve) dagegen noch mit 30 % ihrer maximalen Leistung. Wachstum (blaue Kreise) wurde an Hand der Verdopplungsrate von Kohorten von Zellen gemessen(nach Körner C (2003) Alpine Plant Life. Springer, Berlin).
Nach dem alten Paradigma würde die Photosynthese in der Kälte zum Wachstums-limitierenden Prozess. Tatsächlich reduziert Kälte die Aktivität der Senke, sodass die Rohstoffe der Photosynthese nicht mehr in die Neubildung von Geweben eingesetzt werden können.
Experimentell untersucht: Wie wirkt sich ein Mehr an CO2 auf das Ökosystem Wald aus?
Wälder speichern den überwiegenden Anteil (> 80 %) des in der Biomasse gebundenen Kohlenstoffs unserer Biosphäre. Welchen Effekt ein erhöhtes CO2-Angebot auf das Wachstum der Bäume hat , wurde bisher im wesentlichen nur unter "gestörten Bedingungen untersucht: auf jungen Böden mit ausreichend Nährstoffen, an jungen Bäumen, die oft ohne Konkurrenz zu Nachbarn aufwuchsen. Unter diesen Bedingungen- einem nicht limitierendem Angebot an Ressourcen - kann CO2 einen stimulierenden Effekt von auf das Wachstum haben.
Welchen Effekt hat ein erhöhtes CO2-Angebot aber unter realen Wald-Bedingungen?
In einem bis jetzt einzigartigem Versuchsdesign ("Swiss Canopy Crane Project") haben wir in einem naturbelassenen, artenreichen Mischwald bei Basel das CO2 der Luft rund um die Wipfel von 110 Jahre alten Bäumen auf einen Stand angereichert, der dem voraussichtlichen CO2-Pegel des Jahres 2080 entspricht (530 ppm). Die ausgewählten Bäume waren bis zu 40 m hoch und die CO2 Anreicherung der Luft erfolgte über poröse, feine Schläuche, die von einem 50 m hohen Baukran aus in die Kronen der Bäume (rund 1 km Schlauch pro Baum) gelegt wurden. Abbildung 6.
Das stabile 13C-Isotop als Indikator
Das Experiment lief über 8 Jahre, in denen wir täglich 2 Tonnen CO2 verströmten. Es handelte sich dabei um ein kommerziell erhältliches, auf Lebensmitteltauglichkeit gereinigtes Abfallprodukt der Industrie. Eine Besonderheit dieses CO2 liegt darin, dass es im Vergleich zum atmosphärischen CO2 ein unterschiedliches Verhältnis der beiden natürlich vorkommenden, stabilen Isotope des Kohlenstoffs – 12C und 13C - aufweist: kommen in der Atmosphäre 13C : 12C im Verhältnis von rund 1 : 99 vor, so enthält das aus fossilen Pflanzenresten (Erdöl, Kohle) stammende CO2 Gas etwas weniger 13C. Nach Aufnahme des angereicherten CO2-Gemisches über die Blätter lässt sich die veränderte Isotopensignatur in den pflanzlichen Geweben mittels Massenspektrometrie bestimmen. Damit kann nicht nur bewiesen werden, dass das "neue" CO2 assimiliert wird, sondern, dass - und auch wie rasch - das 13C -Signal tatsächlich in alle Teile des Baumes sickert, in den hölzernen Stamm (Abbildung 6), ebenso wie in die Wurzeln. Der gesamte Weg von der Photosynthese bis zur Verteilung ihrer Produkte und schließlich der Freisetzung im Boden kann so verfolgt werden.
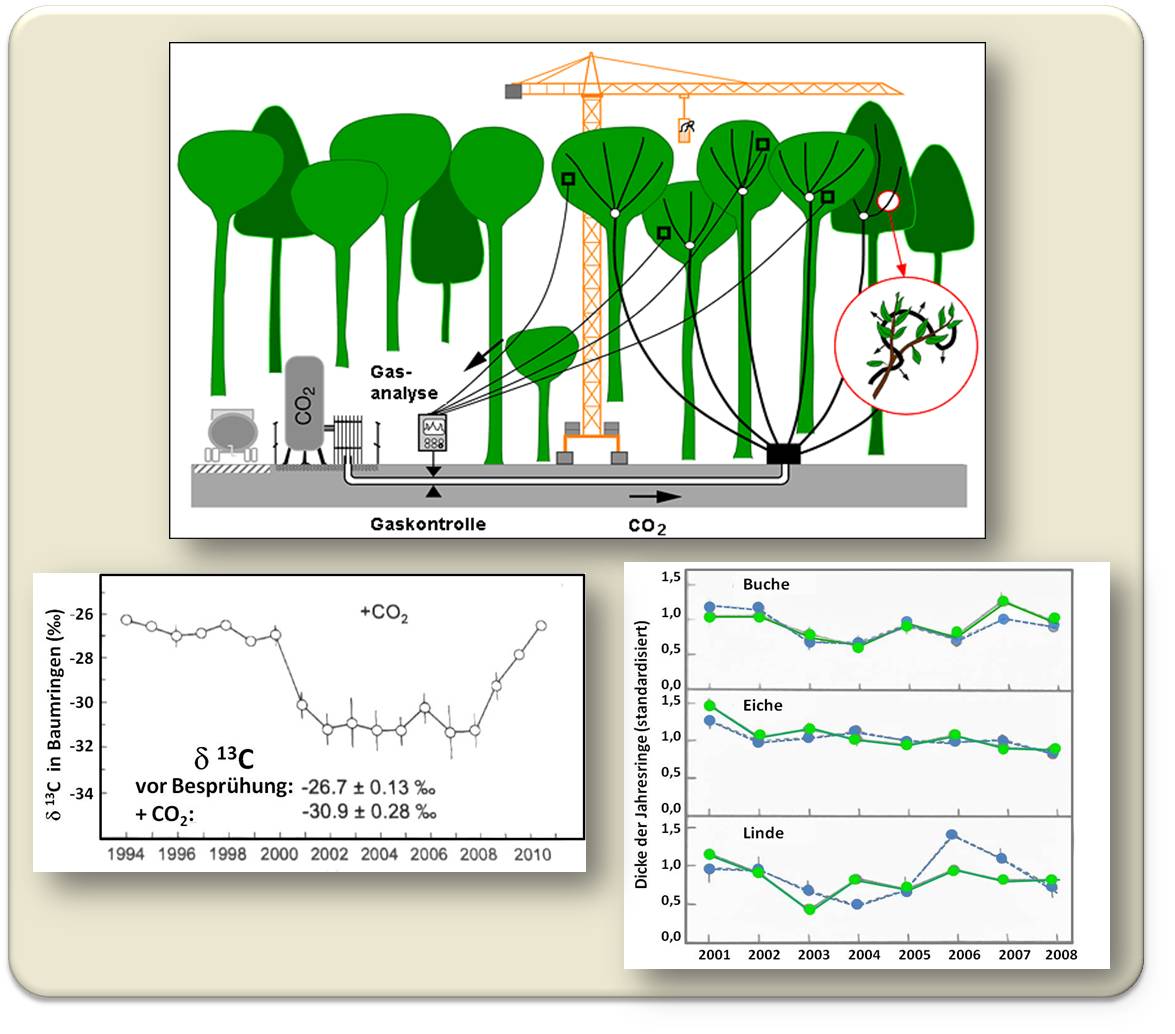 Abbildung 6. Das Swiss Canopy Crane Project: Experimentelle Anreicherung von CO2 in den Kronen hoher Bäume in einem naturbelassenen Mischwald. Oben: Schematische Versuchsanordnung. Die Besprühung mit CO2 erfolgt über poröse Schläuche, die von einem hohen Kran aus in die Kronen der Bäume verlegt wurden, Unten links: Nach Beginn der Besprühung (im Jahr 2000) sinkt der relative 13C-Gehalt in in den Baumringen auf einen konstanten Level, um nach Ende der Behandlung (2008) wieder auf den Anfangswert zu steigen. Unten rechts: Die Dicke der Jahresringe ist unabhängig davon ob Bäume mit CO2 besprüht wurden (grün) oder unbehandelt blieben (blau). (C. Körner et al. 2005 Science 309:1360-1362 und unveröff Daten)
Abbildung 6. Das Swiss Canopy Crane Project: Experimentelle Anreicherung von CO2 in den Kronen hoher Bäume in einem naturbelassenen Mischwald. Oben: Schematische Versuchsanordnung. Die Besprühung mit CO2 erfolgt über poröse Schläuche, die von einem hohen Kran aus in die Kronen der Bäume verlegt wurden, Unten links: Nach Beginn der Besprühung (im Jahr 2000) sinkt der relative 13C-Gehalt in in den Baumringen auf einen konstanten Level, um nach Ende der Behandlung (2008) wieder auf den Anfangswert zu steigen. Unten rechts: Die Dicke der Jahresringe ist unabhängig davon ob Bäume mit CO2 besprüht wurden (grün) oder unbehandelt blieben (blau). (C. Körner et al. 2005 Science 309:1360-1362 und unveröff Daten)
Die Produktivität der Bäume
– von Laubbäumen ebenso wie von Nadelbäumen – wurde mittels mehrerer Methoden bestimmt: u.a. durch Quantifizierung des jeweiligen gesamten Abfalls – Laub oder Nadeln, Zapfen, etc. – oder auch durch Messung der Breite der Jahresringe (Abbildung 6). Weder das Ausmaß des Abfalls noch die Breite der Jahresringe ließen einen Effekt der CO2–Anreicherung auf das Wachstum erkennen . Wir haben solche Versuche auch in anderen Lebensräumen, so zum Beispiel in alpinen Rasen durchgeführt - mit dem gleichen Ergebnis.
Es gibt also keinen Hinweis darauf, dass in naturbelassenen Ökosystemen CO2 in seinem gegenwärtigen Pegel eine für das Wachstum limitierende Ressource darstellt. Ein Mehr an atmosphärischem CO2 wirkt sich daher nicht wie ein Dünger auf das Pflanzenwachstum aus.
Austausch von Kohlenstoff zwischen den Bäumen
Innerhalb von 3 Monaten nachdem wir die CO2-Begasung der Baumkronen begonnen hatten, war das 13C-Signal bereits in den im Umfeld wachsenden Pilzen angelangt. Es war über die feinen Wurzeln der besprühten Bäume dorthin gelangt. Ein völlig unerwartetes Ergebnis war aber, dass sich das Signal auch in den Wurzeln der umliegenden, unbehandelten Bäume ausbreitete. Es hatte offensichtlich den Weg über die verzweigten Myzelien der in Symbiose lebenden Mycorrhizapilze - Täublinge, Schleierlinge, Milchlinge und Ritterlinge - genommen. Rund 40 % der Photosyntheseprodukte in den Wurzeln eines Baums waren so in das Wurzelgeflecht des Nachbarn übergetreten (Klein et al. 2016, Science 352:342-344).
Dies eröffnet eine neue Dimension in dem komplexen Ökosystem Wald: Bäume konkurrieren nicht nur um Licht, Wasser und andere Ressourcen, sie treiben auch ausgedehnten Handel von Baum zu Baum mit den Produkten der Photosynthese. In Hinblick auf den Kohlenstoffhaushalt – Input versus Output – kann ein Baum also nicht mehr als unabhängiges Einzelwesen behandelt werden.
Fazit
Der Kohlenstoffkreislauf wird vom Nährstoffkreislauf und anderen das Pflanzenwachstum begrenzende Bedingungen gesteuert. Die am meisten mangelnde Ressource und die am stärksten limitierenden Umweltbedingungen kontrollieren primär die Prozesse der Gewebeneubildung und diese wiederum bestimmen den Bedarf an Photosyntheseprodukten und damit die Assimilation von CO2.
Erhöhtes CO2 kann das Pflanzenwachstum nur dann steigern, wenn auch ausreichend Nährstoffe zur Verfügung stehen. Wie in Untersuchungen über die Auswirkungen von angereichertem CO2 auf das Pflanzenwachstum gezeigt wurde, ist dies in naturbelassenen Wäldern kaum der Fall. Dass erhöhtes CO2 in der Atmosphäre zu einem nachhaltigen Anstieg der Kohlenstoffspeicherung in der Biomasse führt, ist demnach unwahrscheinlich .
Weiterführende Links
Webseite von Christian Körner: https://plantecology.unibas.ch/koerner/index.shtml
Der Wald in einer CO2-reichen Welt http://www.waldwissen.net/wald/klima/wandel_co2/wsl_wald_co2/index_DE (abgerufen am 19.7.2016)
Interview mit Christian Körner (28.10.2014) http://www.basellandschaftlichezeitung.ch/basel/basel-stadt/die-pflanzen... (abgerufen am 19.7.2016)
Kohlenstoffhandel von Baum zu Baum (15.4.2016) http://www.arboristik.de/baumpflege_wissen_15042016.html (abgerufen am 19.7.2016)
Swiss Plant Science Web: Ecosystem function and biodiversity under global change. https://swissplantscienceweb.ch/nc/research/home/portfolio/koerner/ (abgerufen am 19.7.2016)
Videos (in Englisch)
Christian Körner: On Plants and Carbon. Special Lecture in commemoration of Professor Kurt Komarek (6.6.2016). 43:12 min. https://www.youtube.com/watch?v=i56VcHoBvjE
Christian Körner: Is the biosphere carbon limited? (23.6.2015). 34:14 min. https://www.youtube.com/watch?v=Cav2czzjDFQ
Christian Körner: What carbon cyclists can learn from bankers (BES Science Slam 2015). 10:37 min. British Ecological Society https://www.youtube.com/watch?v=O5kQHw23VbY
Artikel zum Themenkomplex Kohlenstoffkreislauf - Kohlenstoffspeicherung im Scienceblog
Redaktion, 26.06.2015: Die Erde ist ein großes chemisches Laboratorium – wie Gustav Tschermak vor 150 Jahren den Kohlenstoffkreislauf beschrieb
Walter Kutschera, 22.01.2016: Radiokohlenstoff als Indikator für Umweltveränderungen im Anthropozän
Rattan Lal, 27.11.2015: Boden - Der große Kohlenstoffspeicher
Rattan Lal, 11.12.2015: Der Boden – die Lösung globaler Probleme liegt unter unseren Füßen
Knut Ehlers, 01.04.2016: Der Boden - ein unsichtbares Ökosystem
Rupert Seidl, 18.03.2016: Störungen und Resilienz von Waldökosystemen im Klimawandel
Julia Pongratz & Christian Reick, 18.07.2014: Landwirtschaft pflügt das Klima um
Gerhard Glatzel, 21.03.2013: Rückkehr zur Energie aus dem Wald — mehr als ein Holzweg? (Teil 1)
Gerhard Glatzel, 04.04.2013: Rückkehr zur Energie aus dem Wald — mehr als ein Holzweg? (Teil 2)
Antje Boetius, 13.05.2016: Mikrobiome extremer Tiefsee-Lebensräume
Christa Schleper, 19.06.2015: Erste Zwischenstufe in der Evolution von einfachsten zu höheren Lebewesen entdeckt: Lokiarchaea
Gerhard Herndl, 21.10.2014: Das mikrobielle Leben der Tiefsee
Gottfried Schatz, 27.09.2012: Sonnenkinder — Wie das atomare Feuer der Sonne die Meerestiefen erhellt
Gottfried Schatz, 23.02.2012: Erdfieber — Das Unbehagen der Wissenschaft bei der Klimadebatte
- Printer-friendly version
- Log in to post comments