Die biomimetische künstliche Nase – wie weit sind wir? Teil 2. Aufbau und Funktion physiologischer Geruchssensoren
Die biomimetische künstliche Nase – wie weit sind wir? Teil 2. Aufbau und Funktion physiologischer GeruchssensorenFr, 26.01.2012- 04:20 — Wolfgang Knoll
![]()

Fortsetzung von Teil 1: Künstliche Sensoren nach dem natürlichen Vorbild unserer fünf Sinne, erschienen am 12.Jänner 2012).Einige heute bekannte Details über den Aufbau der Geruchssensoren, und zwar sowohl für Wirbeltiere als auch für Wirbellose, also z.B. den Insekten, sind stark vereinfacht in Abbildung 3 gegeben.
Bei den Wirbeltieren, also auch bei der Ratte, dem Hund und beim Menschen befinden sich die meisten Nervenzellen im Riech-Epithel (Riechschleimhaut) im Dach der Nasenhaupthöhle. Hier sitzen Millionen von Riechzellen. Die Signale werden von dort über den Riechnerv direkt an das Gehirn weitergeleitet. Die Riechzellen (Olfactory Sensory Neurons) reichen mit ihren Riechhaaren (Ciliae) bis in die Nasenschleimhaut (Mucosa), die mit ihrem Sekret (Mucus) die Zellen und ihre Membranen vor dem Austrocknen schützen müssen, da diese im direkten Kontakt mit der eingeatmeten Luft mit den mitgeführten zu detektierenden Duftstoffen und Pheromonen steht. 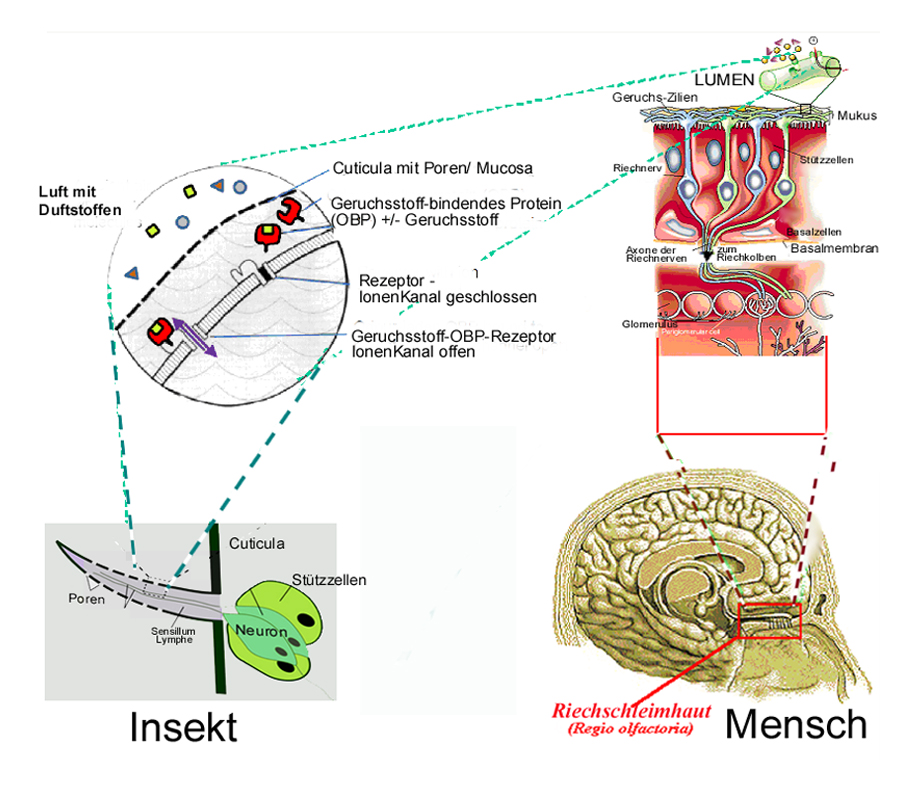
Abbildung 3: Zum Aufbau der Geruchssensoren von Wirbeltieren (Mensch, rechte Bilder) und Wirbellosen (Insekten, linker Cartoon) mit dem zentralen Element einer durch Geruchs- Rezeptoren funktionalisierten Membran im Zentrum (im Kreis).
Bei Wirbellosen wird der Riechnerv in den Sensillen der Antennen durch die sogenannte Cuticula (selbsttragende „Körperdecke“) mechanisch geschützt und vor dem Austrocknen bewahrt. Die durch die vorbei streichende Luft antransportierten Duftstoffe können durch Poren in der Cuticula den Riechnerv erreichen, welcher von der Lymphe umgeben ist und bei Bindung eines Duftstoffes oder eines (Art-) spezifischen Pheromons ein bestimmtes elektrisches Signal, die Spikes, generiert.
Wie erreichen Duftstoffe ihre Geruchssensoren?
Da die über Geruchssensoren ablaufenden molekularen Prozesse die Basis für jede Überlegung zum Konzept und Bau einer künstlichen biomimetischen Nase darstellen, sollen sie im Folgenden noch etwas genauer, wenn auch nach wie vor sehr schematisch dargestellt werden. Dazu betrachten wir die Abbildung 4: 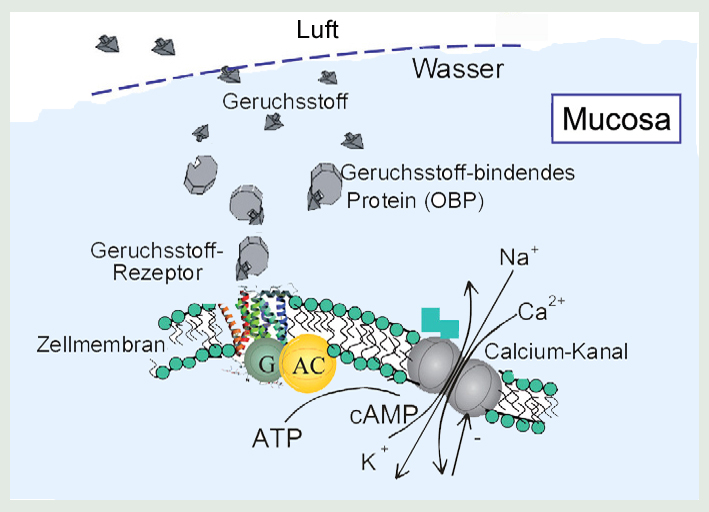
Abbildung 4: Die wesentlichen molekularen Einzelschritte, die bei Erkennung und Bindung eines Duftstoffes (Odorants) zu einem elektrischen Signal führen. Geruchsstoffrezeptor ist ein G-Protein gekoppelter Rezeptor (GPCR), G: G-Protein, AC: Adenylat-Cyclase
Geruchsstoffe (Odorants) aus der Luft müssen an irgendeiner Stelle den Übergang in die wässrige Phase schaffen. Bei den Wirbellosen passiert dies beim Durchtritt durch die Poren in der Cuticula, während der in Abbildung 4 dargestellte Fall eher typisch für Wirbeltiere ist, wo dieser Übergang in der Grenzfläche der Mucosa zur Luft geschieht. Da es sich bei vielen Duftstoffen um kleine und vor allem hydrophobe (d.h. wasserabweisende) Moleküle handelt, sorgen die Geruchsstoff-bindenden Proteine (Odorant Binding Proteins - OBPs) oder Pheromon-bindende Proteine (PBPs), welche mit den Duftstoffen einen Komplex bilden, nicht nur für eine erhöhte Löslichkeit, sie „shutteln“ die Duftstoffe zu den in der Membran sitzenden Duftstoff-Rezeptoren (Odorant Receptors, ORs). Ob dies allein durch freie Diffusion geht ist noch unklar; bei den Wirbellosen wird wegen der hohen OBP Konzentration in der Lymphe auch ein gerichteter Transport entlang perkolierender Protein- Aggregate diskutiert.
Bindung von Duftstoffen an den Rezeptor und Signaltransfer
Weiterhin auch nicht völlig klar ist, ob die OBPs auch an der entscheidenden Bindung der Duftstoffe an den Rezeptor beteiligt sind oder einfach nur ihr Cargo abgeben. Als gesichert gilt aber, dass die Bindung der Odorants eher weniger spezifisch ist, allerdings die der Pheromone an ihre entsprechenden hochspezifischen Rezeptoren, eine Reaktionskaskade auslöst, die die wesentlichen Verstärkungsmechanismen für die hohe Empfindlichkeit beim Riechen ausmacht. Hier sollte man darauf hinweisen, dass die gegenüber dem Menschen deutlich höhere „Riechleistung“ des Hundes nicht auf einem anderen Mechanismus beruht, sondern lediglich vor allem Ausdruck der Tatsache ist, dass wir Menschen in Summe nur etwa 5 cm2 aktive Fläche mit Riechzellen haben, während der Hund auf das Fünffache kommt. Aber auch beim Weltmeister der Riechchampions, dem Seidenspinner Bombyx Mori, der nur ganz wenige Moleküle des von den Weibchen ausgesandten Pheromones Bombycol benötigt, um die Angebetete zu finden, sind die molekularen Mechanismen der Duftstofferkennung sehr ähnlich.
Die Bindung von Odorants an der einen Seite der Membran an transmembrane, also Membran-überspannende ORs, die alle zur großen Klasse der G-Protein gekoppelten Rezeptoren (G-Protein Coupled Receptors, GPCRs) gehören, führt auf der anderen Seite der Membran zur Dissoziation des G-Proteins als Trigger des Signal-Übertragungsmechanismus.* Die ?-Untereinheit de G-Proteins bindet an die Adenylat-Cyclase, einem integralen Membran-Enzym, das aus Adenosintriphosphat (ATP) die Synthese eines sogenannten second Messenger – des cyclischen Adenosine Monophosphat (cAMP) - katalysiert. Da die Bindung eines einzigen Duftmoleküls zur Synthese von über 1000 cAMP Molekülen führt, ist damit ein erster Verstärkungsmechanismus identifiziert.
In weiterer Vereinfachung der biologischen Komplexität fasst dann Abb. 4 zusammen, dass der second messenger cAMP Ionen-Kanäle in ihrer Permeabilität beeinflusst: so wirkt das cAMP direkt auf einen Ionen-Kanal, der bei Bindung von einem cAMP Molekül geöffnet wird. Durch diesen Kanal passieren dann zwischen einer Million und 100 Millionen Calcium- und Natrium-Ionen die Membran von außen nach innen, während Kalium-Ionen ausströmen können – eine weitere enorme Verstärkung eines einzelnen molekularen Ereignisses. Die damit verbundenen transienten, elektrischen Signale sind die Basis der Informationsverarbeitung beim Riechen in den Glomeruli und dann letztendlich im Gehirn.
Anmerkungen der Redaktion
Aufbau und Funktion von G-Protein Coupled Receptors (GPCRs) und G-Proteinen sind beschrieben in dem einleitenden Artikel „Wie wir die Welt um uns herum wahrnehmen“ (erschienen am 5.1.2011)
Glossar
Adenosintriphosphat (ATP): Universeller Energielieferant für zelluläre Prozesse. Bei (enzymatischer) Spaltung der Phosphatbindungen wird Energie freigesetzt, die u.a. zur Synthese von Biomolekülen, zum Stofftransport durch Membranen, zur Muskeltätigkeit,.. benötigt wird. Daneben spielt ATP auch eine bedeutende Rolle im Signaltransfer.
Adenylatcyclase: katalysiert die Bildung des second messenger cyclo-AMP (c-AMP) aus ATP unter Abspaltung des Pyrophosphatrestes.
Glomeruli: kugelige Gebilde im Riechkolben in denen sich die Nerven aller Riechzellen sammeln (jeweils ein Glomerulus je Typ Riechzelle); Abb. 3.
Pheromone: Flüchtige Substanzen, die Information zwischen Mitgliedern derselben Spezies vermitteln. Sie werden von einem Individuum sezerniert und wirken auf das Verhalten eines anderen Individuums
Second Messenger: sekundärer Botenstoff; intrazelluläre Verbindung, deren Konzentration sich als Antwort auf ein von außen kommendes primäres Signal (ausgelöst z.B. durch Andocken eines Duftstoffs am Rezeptor) ändert und, die damit ein von außen kommendes Signal in der Zelle weiterleitet.